ID:SRC_HUMAN DESCRIPTION: RecName: Full=Proto-oncogene tyrosine-protein kinase Src; EC=2.7.10.2; AltName: Full=Proto-oncogene c-Src; AltName: Full=pp60c-src; Short=p60-Src; FUNCTION: Non-receptor protein tyrosine kinase which is activated following engagement of many different classes of cellular receptors including immune response receptors, integrins and other adhesion receptors, receptor protein tyrosine kinases, G protein- coupled receptors as well as cytokine receptors. Participates in signaling pathways that control a diverse spectrum of biological activities including gene transcription, immune response, cell adhesion, cell cycle progression, apoptosis, migration, and transformation. Due to functional redundancy between members of the SRC kinase family, identification of the specific role of each SRC kinase is very difficult. SRC appears to be one of the primary kinases activated following engagement of receptors and plays a role in the activation of other protein tyrosine kinase (PTK) families. Receptor clustering or dimerization leads to recruitment of SRC to the receptor complexes where it phosphorylates the tyrosine residues within the receptor cytoplasmic domains. Plays an important role in the regulation of cytoskeletal organization through phosphorylation of specific substrates such as AFAP1. Phosphorylation of AFAP1 allows the SRC SH2 domain to bind AFAP1 and to localize to actin filaments. Cytoskeletal reorganization is also controlled through the phosphorylation of cortactin (CTTN). When cells adhere via focal adhesions to the extracellular matrix, signals are transmitted by integrins into the cell resulting in tyrosine phosphorylation of a number of focal adhesion proteins, including PTK2/FAK1 and paxillin (PXN). In addition to phosphorylating focal adhesion proteins, SRC is also active at the sites of cell-cell contact adherens junctions and phosphorylates substrates such as beta-catenin (CTNNB1), delta-catenin (CTNND1), and plakoglobin (JUP). Another type of cell-cell junction, the gap junction, is also a target for SRC, which phosphorylates connexin- 43 (GJA1). SRC is implicated in regulation of pre-mRNA-processing and phosphorylates RNA-binding proteins such as KHDRBS1. Also plays a role in PDGF-mediated tyrosine phosphorylation of both STAT1 and STAT3, leading to increased DNA binding activity of these transcription factors. Involved in the RAS pathway through phosphorylation of RASA1 and RASGRF1. Plays a role in EGF-mediated calcium-activated chloride channel activation. Required for epidermal growth factor receptor (EGFR) internalization through phosphorylation of clathrin heavy chain (CLTC and CLTCL1) at 'Tyr- 1477'. Involved in beta-arrestin (ARRB1 and ARRB2) desensitization through phosphorylation and activation of ADRBK1, leading to beta- arrestin phosphorylation and internalization. Has a critical role in the stimulation of the CDK20/MAPK3 mitogen-activated protein kinase cascade by epidermal growth factor. Might be involved not only in mediating the transduction of mitogenic signals at the level of the plasma membrane but also in controlling progression through the cell cycle via interaction with regulatory proteins in the nucleus. Plays an important role in osteoclastic bone resorption in conjunction with PTK2B/PYK2. Both the formation of a SRC-PTK2B/PYK2 complex and SRC kinase activity are necessary for this function. Recruited to activated integrins by PTK2B/PYK2, thereby phosphorylating CBL, which in turn induces the activation and recruitment of phosphatidylinositol 3-kinase to the cell membrane in a signaling pathway that is critical for osteoclast function. Promotes energy production in osteoclasts by activating mitochondrial cytochrome C oxidase. Phosphorylates DDR2 on tyrosine residues, thereby promoting its subsequent autophosphorylation. Phosphorylates RUNX3 and COX2 on tyrosine residues, TNK2 on 'Tyr-284' and CBL on 'Tyr-731'. Enhances DDX58/RIG-I-elicited antiviral signaling. Phosphorylates PDPK1 at 'Tyr-9', 'Tyr-373' and 'Tyr-376'. CATALYTIC ACTIVITY: ATP + a [protein]-L-tyrosine = ADP + a [protein]-L-tyrosine phosphate. ENZYME REGULATION: Phosphorylation by CSK at Tyr-530 inhibits kinase activity. Inhibitory phosphorylation at Tyr-530 is enhanced by heme. Further phosphorylation by CDK1 partially reactivates CSK-inactivated SRC and facilitates complete reactivation by protein tyrosine phosphatase PTPRC. Integrin engagement stimulates kinase activity. Phosphorylation by PTK2/FAK1 enhances kinase activity. Butein and pseudosubstrate-based peptide inhibitors like CIYKYYF act as inhibitors. Phosphorylation at Tyr-419 increases kinase activity. SUBUNIT: Interacts with DDEF1/ASAP1; via the SH3 domain. Interacts with CCPG1. Identified in a complex containing FGFR4, NCAM1, CDH2, PLCG1, FRS2, SRC, SHC1, GAP43 and CTTN. Interacts with ERBB2, STAT1 and PNN. Interacts with DDR1, DDR2 and DAB2. Interacts with CDCP1, PELP1, TGFB1I1 and TOM1L2. Interacts with the cytoplasmic domain of MUC1, phosphorylates it and increases binding of MUC1 with beta-catenin. Interacts with RALGPS1; via the SH3 domain. Interacts with HEV ORF3 protein; via the SH3 domain. Interacts with CAV2 (tyrosine phosphorylated form). Interacts (via the SH3 domain and the protein kinase domain) with ARRB1; the interaction is independent of the phosphorylation state of SRC C-terminus. Interacts with ARRB1 and ARRB2. Interacts with SRCIN1. Interacts with NDFIP2 and more weakly with NDFIP1. Interacts with PIK3CA and/or PIK3C2B, PTK2/FAK1 and ESR1 (dimethylated on arginine). Interacts with FASLG. Interacts (via SH2 domain) with the 'Tyr- 402' phosphorylated form of PTK2B/PYK2. Interacts (via SH2 domain) with FLT3 (tyrosine phosphorylated). Interacts with PDGFRA (tyrosine phosphorylated). Interacts with CSF1R. Interacts (via SH2 and SH3 domain) with TNK2. Interacts (via protein kinase domin) with the tyrosine phosphorylated form of RUNX3 (via runt domain). Interacts with TRAF3 (via RING-type zinc finger domain). Interacts with DDX58, MAVS and TBK1. Interacts (via SH2 domain) with GNB2L1/RACK1; the interaction is enhanced by tyrosine phosphorylation of GNB2L1 and inhibits SRC activity. Interacts with EPHB1; activates the MAPK/ERK cascade to regulate cell migration. Interacts with FCAMR. Interacts (via SH2 domain) with the 'Tyr-9' phosphorylated form of PDPK1. Interacts with AMOTL2; this interaction regulates the translocation of phosphorylated SRC to peripheral cell-matrix adhesion sites. INTERACTION: P42684:ABL2; NbExp=2; IntAct=EBI-621482, EBI-1102694; P12814:ACTN1; NbExp=2; IntAct=EBI-621482, EBI-351710; Q8R5G7:Arap3 (xeno); NbExp=3; IntAct=EBI-621482, EBI-621463; Q9ULH1:ASAP1; NbExp=2; IntAct=EBI-621482, EBI-346622; P12830:CDH1; NbExp=2; IntAct=EBI-621482, EBI-727477; P00533:EGFR; NbExp=5; IntAct=EBI-621482, EBI-297353; P04626:ERBB2; NbExp=10; IntAct=EBI-621482, EBI-641062; P03372-4:ESR1; NbExp=2; IntAct=EBI-621482, EBI-4309277; P25445:FAS; NbExp=2; IntAct=EBI-621482, EBI-494743; Q9Y6K9:IKBKG; NbExp=3; IntAct=EBI-621482, EBI-81279; P35968:KDR; NbExp=2; IntAct=EBI-621482, EBI-1005487; Q07666:KHDRBS1; NbExp=3; IntAct=EBI-621482, EBI-1364; Q05397:PTK2; NbExp=4; IntAct=EBI-621482, EBI-702142; Q13905:RAPGEF1; NbExp=2; IntAct=EBI-621482, EBI-976876; Q01973:ROR1; NbExp=9; IntAct=EBI-621482, EBI-6082337; Q9C0H9:SRCIN1; NbExp=3; IntAct=EBI-621482, EBI-1393949; Q68CZ2:TNS3; NbExp=13; IntAct=EBI-621482, EBI-1220488; SUBCELLULAR LOCATION: Cell membrane. Mitochondrion inner membrane. Nucleus. Cytoplasm, cytoskeleton. Note=Localizes to focal adhesion sites following integrin engagement. Localization to focal adhesion sites requires myristoylation and the SH3 domain. TISSUE SPECIFICITY: Expressed ubiquitously. Platelets, neurons and osteoclasts express 5-fold to 200-fold higher levels than most other tissues. DOMAIN: The SH2 and SH3 domains are important for the intramolecular and intermolecular interactions that regulate catalytic activity, localization, and substrate recruitment. PTM: Myristoylated at Gly-2, and this is essential for targeting to membranes. PTM: Dephosphorylated at Tyr-530 by PTPRJ (By similarity). Phosphorylated on Tyr-530 by c-Src kinase (CSK). The phosphorylated form is termed pp60c-src. Dephosphorylated by PTPRJ at Tyr-419. Normally maintained in an inactive conformation with the SH2 domain engaged with Tyr-530, the SH3 domain engaged with the SH2-kinase linker, and Tyr-419 dephosphorylated. Dephosphorylation of Tyr-530 as a result of protein tyrosine phosphatase (PTP) action disrupts the intramolecular interaction between the SH2 domain and Tyr-530, Tyr-419 can then become autophosphorylated, resulting in SRC activation. Phosphorylation of Tyr-530 by CSK allows this interaction to reform, resulting in SRC inactivation. CDK5-mediated phosphorylation at Ser-75 targets SRC to ubiquitin-dependent degradation and thus leads to cytoskeletal reorganization. Phosphorylated by PTK2/FAK1; this enhances kinase activity. Phosphorylated by PTK2B/PYK2; this enhances kinase activity. PTM: S-nitrosylation is important for activation of its kinase activity (By similarity). PTM: Ubiquitinated in response to CDK5-mediated phosphorylation. DISEASE: Note=SRC kinase activity has been shown to be increased in several tumor tissues and tumor cell lines such as colon carcinoma cells. SIMILARITY: Belongs to the protein kinase superfamily. Tyr protein kinase family. SRC subfamily. SIMILARITY: Contains 1 protein kinase domain. SIMILARITY: Contains 1 SH2 domain. SIMILARITY: Contains 1 SH3 domain.
The RNAfold program from the Vienna RNA Package is used to perform the secondary structure predictions and folding calculations. The estimated folding energy is in kcal/mol. The more negative the energy, the more secondary structure the RNA is likely to have.
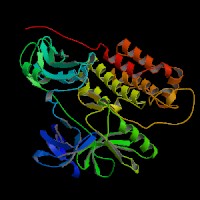
ModBase Predicted Comparative 3D Structure on P12931
Front
Top
Side
The pictures above may be empty if there is no ModBase structure for the protein. The ModBase structure frequently covers just a fragment of the protein. You may be asked to log onto ModBase the first time you click on the pictures. It is simplest after logging in to just click on the picture again to get to the specific info on that model.
Orthologous Genes in Other Species
Orthologies between human, mouse, and rat are computed by taking the best BLASTP hit, and filtering out non-syntenic hits. For more distant species reciprocal-best BLASTP hits are used. Note that the absence of an ortholog in the table below may reflect incomplete annotations in the other species rather than a true absence of the orthologous gene.
 Sequence and Links to Tools and Databases
Sequence and Links to Tools and Databases  Common Gene Haplotype Alleles
Common Gene Haplotype Alleles